Soil biology revealed
Publication review by Patrick Francis
Soil biology is becoming the world’s most engaging scientific frontier. It can claim that title because the organisms involved and their activity are critical for life on earth but scientific knowledge of its holistic impacts is limited.
Up until soil biologists such as Dr Elaine Ingham published “The Soil Food Web” (a USDA book and e-book) the world of soil biology was rarely related to soil health, nutrition and structure. Agricultural science seldom mentioned soil health supporting organisms except for one key class of bacteria (rhizobia) and fungi (mycorrhizae) which form symbiotic relationships with legume and grass plants respectively.
Over the last two decades the soil biology world has become recognised as the most critically important component in rectifying, maintaining and improving soil health with all its associated ecosystem services. And soil biology populations are intricately associated with the levels of soil organic matter (SOM) and soil organic carbon (SOC).

Figure: “One teaspoon of soil contains more living organisms than there are people in the world”. Source: State of Knowledge of Soil Biodiversity, FAO Report 2020.
The problem with this FAO statement is that soils and environments are so different at the local and world scale it is virtually meaningless.
Just where scientific soil biology knowledge has reached is superbly documented in the Food and Agriculture Organisation (FAO) of the United Nations 2020 Report “State of knowledge of soil biodiversity – Status, challenges and potentialities”. It is chapters 2 and 3 which make compelling reading for anyone involved in land management and agriculture anywhere in the world.
Given that preventing climate change is the world’s number one environmental and social issue there is one section of this report which stands out. It’s the discussion around interactions between organisms and the soil food web. The graphic and caption highlights what few people understand, that the soil food web is one of the major contributors to greenhouse gas emissions. Ironically the Report does not reflect on this and fails to point out that the emissions contribute to greenhouse gas flows, not atmospheric stocks as generated by burning fossil fuels.
What the report fails to point out is that increasing soil health and building soil carbon is associated with increasing soil biota and subsequently emissions. In a warming climate which further enhances soil biology it is difficult to understand how soil carbon has a significant role to play in carbon abatement unless it is being undertaken on soils previously degraded by farming or mining.
The Report summarises the soil food web exceptionally well: “Through photosynthesis, plants transform light energy into chemical energy, allowing them to produce their own organic compounds -carbohydrates- to live. Plant and microbial activity account for 90 % of the energy transfer in terrestrial ecosystems. Most carbon and nutrients are transferred to the soil via litterfall, root exudation and decaying organisms (and manures). Bacteria, fungi and archaea that inhabit the root microbiome or are root symbionts benefit from this input. Plants return about half of this carbon to the atmosphere through respiration in the form of carbon dioxide (CO2).
“Different microorganisms return carbon and nitrogen to the atmosphere, making the continuity of life possible. Most carbon entering the soil is lost through microbial respiration in the form of CO2. A broad array of soil denitrifying bacteria produce progressively reduced products ending in N2, which returns to the atmosphere. In the absence of oxygen and on organic substrates, some soil bacteria produce methane (CH4).”
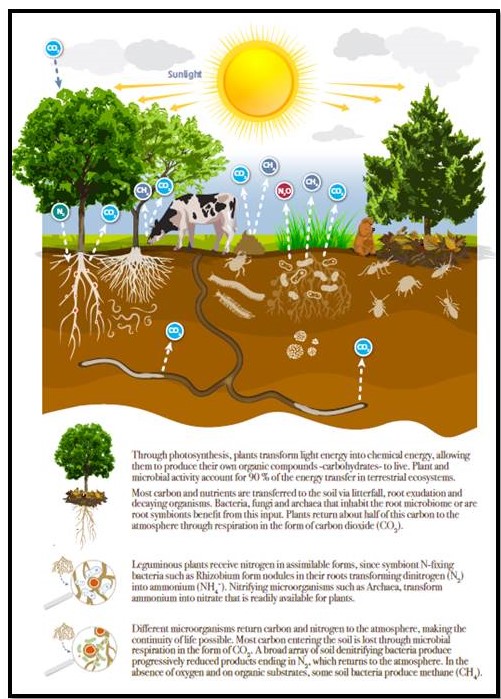
Figure: This outstanding graphic summarises the interactions between plants, livestock and soil food web biota and their roles in greenhouse gases abatement and generation. Source: State of Knowledge of Soil Biodiversity, FAO Report 2020.
Chapter 2 covers “Global diversity and distribution of soil biodiversity” by delving into the roles played by each known biota group. The descriptions and images are mind blowing. I have never read such a detailed science based account of the different species and how each impacts soil and water ecosystem functions. Each sub-chapter describes the impact of particular biota under a range of headings:
* Population description and distribution
* Carbon transformation
* Nutrient cycling
* Soil structure
* Biological regulation
Excerpts from the sub-chapter on Nematodes demonstrates the depth of information provided on each species of soil biota.:
“Nematodes (or roundworms) are the most abundant group of animals in soils and in aquatic systems, representing around 80 percent of all animals on Earth. Nematodes fill a number of trophic roles (A trophic level is the group of organisms within an ecosystem which occupy the same level in a food chain – source: Biology Dictionary) in soils, including as bacterivorous and/ or fungivorous grazers, predators of other nematodes and smaller animals, plant-feeders, omnivores and parasites of both animals and plants. …It is estimated that globally, soil nematodes operate on a monthly C budget of 139 Mt. Of this, nearly 110 Mt C is respired, which is equal to approximately 15 percent of the C released from fossil fuels .

Figure: A soil nematode. Source: State of Knowledge of Soil Biodiversity, FAO Report 2020.
“Nematodes are responsible for the transfer of many nutrients through trophic networks. For example, nematodes retain higher amounts of nitrogen (N) than they need; this is expelled as ammonia, which after nitrification can be readily consumed by plants and bacteria. Similarly, N leaching has been shown to increase with nematode biomass .
“Conversely, grazing of denitrifying bacteria by nematodes has been shown to cause a nearly 8 percent decline in denitrification rates in a mesocosm experiment . The presence of nematodes significantly increases net N (+25 percent) and net P (+23 percent) availability, as well as plant biomass production (+9 percent) compared to their absence . Therefore, free-living soil nematodes are an important component in soil nutrient cycles.
“Direct interactions between soil structure and nematodes are long established and have shown that size of water-filled pores is the major structural limitation for soil nematodes. Nematode communities are often used as a marker of soil quality.
“Many nematode species are parasitic and are important disease agents in crops, livestock and humans. Certain parasitic nematodes have been identified as biocontrol agents of insect populations, especially those with a soil-dwelling larval stage. The use of nematodes as a biocontrol agent has proven popular as they offer an alternative to chemical pesticides. However, there has been little research in understanding the long-term impacts of nematode biocontrols. There are concerns that the global use of nematode biocontrols could lead to unintended killing of non-target insects, especially predators, and may lead to the establishment of invasive nematode populations when species are sold and used outside of their natural range.”
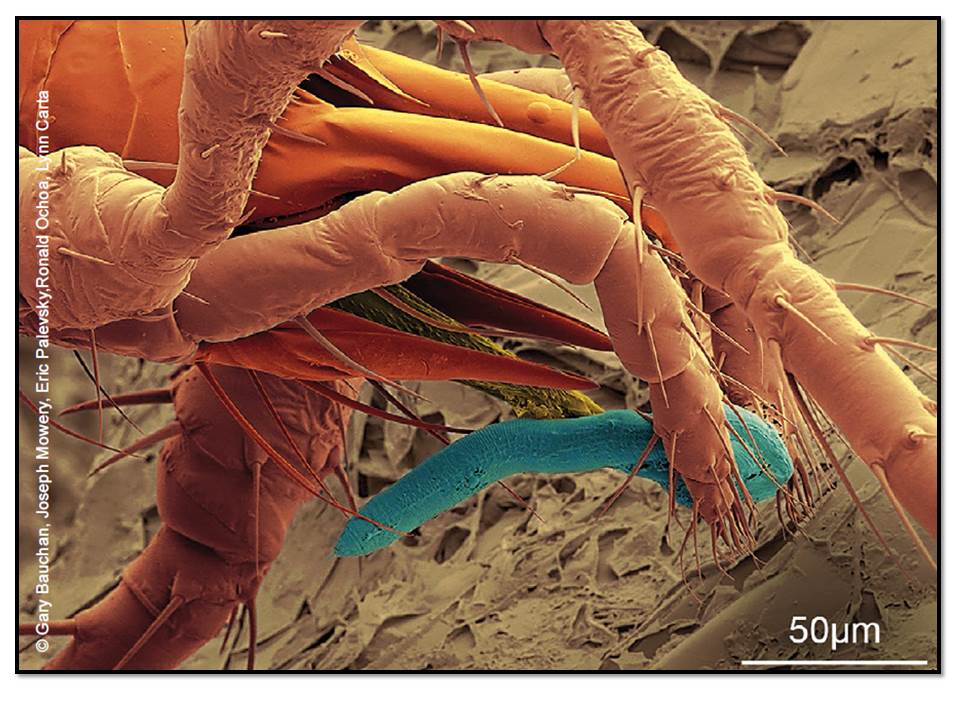
Figure: A mite preys on a nematode. Biota diversity is critical for soil food web balance. Source: State of Knowledge of Soil Biodiversity, FAO Report 2020.
The balance provided in this discussion is particularly important, it helps counter often held perceptions about nature and natural systems – that they are always positive for life and don’t have negative impacts on different life forms in their complex world.
For instance it was revealing to learn that nematodes are responsible for annually releasing into the atmosphere an equivalent of 15% of the carbon released from fossil fuels. Such data highlights the need for caution when applying rules or policies around anthropogenic climate change targets when it comes to agriculture, which to operate successfully must support natures ecosystems functions.
The World Soil Charter (FAO 2015) and the Voluntary Guidelines for Sustainable Soil Management (FAO 2017) emphasise that soil biodiversity provides a wide range of biological functions which are critically important for the future of soil health and world food production.

Figure: Ants are responsible for a range of ecosystem functions through burrowing and soil turnover, through preying on other biota and as being prey for other trophic levels such as Echidnas in Australia. Photos: Patrick Francis.
Related to understanding soil biota’s ecosystem functions impacts is the fact that the science has yet to learn about the number of species involved and how they operate and subsequently what their impacts are. The discussion around soil bacteria states: ”The studies of global microbial diversity … suggest the existence of a potentially extensive group of unclassified bacterial species . These undescribed microorganisms may have a wide range of metabolic capabilities influencing ecosystem services, and may even be of particular interest for specific industrial and environmental management purposes. Better understanding of how biodiversity loss by anthropogenic activities might affect the stability of food webs and ecosystem functioning requires a knowledge of current gaps in soil microbial diversity.”
The unknowns around soil biota are a critical issue surrounding adopting strategies for using species to improve soil health. In such a complex world of interactions between bioata species, plant species, soil types, and rainfall and temperature variation, adding specific species from manufactured teas to produce particular production outcomes would seem to be a long way off. The key being ensuring as wide as possible species diversity, but critical numbers active at different times of the year to produce particular ecosystem function or agronomy outcomes is still mostly unknown.

Figure: Soil food web diversity such as this photo of cattle pasture soil in Victoria, Australia, is a key indicator of biodiversity health. Photo: Patrick Francis
The report notes this issue with respect to soil fungi: “As land use intensifies, there is a dire need for fast and economically viable methods for determining the health status of soil biodiversity. Molecular approaches have developed to the point that technology is no longer the bottleneck, but data is. More research is needed globally to calibrate our knowledge of soil fungal diversity in different biomes, ecosystems and anthropogenic settings. It is only through extensive background knowledge that we begin to understand, manage and conserve soil fungi, soil biodiversity and the functions it provides for humanity.
“While many studies have quantified the effects of agricultural management practices on ecosystem functioning, fewer have linked changes in soil organism diversity directly to changes in ecosystem functioning. …Future research exploring a more direct link between soil biodiversity and ecosystem functioning will improve our understanding of agricultural intensification impacts on biodiversity and ecosystem functioning.”
The report highlights that the most universal characteristic of soil which positively impacts biota mass is soil organic carbon (SOC) percentage (in other words soil organic matter content). Higher SOC supports larger microbial biomass as well as microbial diversity. But global patterns for biota diversity in soils are poorly understood and not always explained by SOC content. It cited an example where research shows that earthworm diversity in Europe is not related to soil organic matter content.
“Nonetheless, the generally positive relationship between soil C stock and soil biodiversity suggests that soil carbon loss is a threat to soil biodiversity.”
Interestingly the report points out that there can be other drivers behind soil biota loss, for instance soil pH. In one European study soil biodiversity was higher in agricultural soil than in carbon rich forest soil with a lower pH. There is also an impact of soil carbon quality (related to its carbon to nitrogen ratio) on biota diversity. One investigation showed that SOC content explained 5% of the variation in bacterial diversity while SOC quality explained 22%
Overall the report concludes that the effects of SOC loss on soil biodiversity are globally poorly understood because:
(i) data on soil biodiversity and the patterns of distribution at large scale are insufficient (especially for soil fauna),
(ii) the biological mechanisms involved in how SOC affects soil biodiversity are poorly known, and
(iii) many of the other threats to soil biodiversity also lead to changes in SOC content, especially land use and climate change.
On the last point about responses of biodiversity to climate change, the report concludes that very little is known and are difficult to predict.
Rest of the report
Chapter 4 covers “Threats to soil biodiversity – global and regional trends”. The threats coverage is interesting but the regional status seems misleading given the coverage of what has been happening in Australia over the past 30 years is anything to go by. It highlights the negatives with no acknowledgement of the enormous steps forward taken by farmers to improve soil health and ecosystem functions through initiatives like:
* Three decades of landcare with ground cover and revegetation high priorities.
* Virtual universal acceptance of conservation cropping, including no till, stubble retention, species rotations, precision farming and variable rate inputs.
* Holistic grazing management and rotational grazing roles in matching livestock paddock stocking rate to paddock livestock carrying capacity to ensure ground cover and ecosystem functions like soil biota are protected and carbon flow into soil is optimised.
* Research to support on-farm improvement of ecosystem functions including protecting and enhancing the soil food web across all farming systems through industry R &D organisations, state departments of agriculture, natural resource management agencies, and CSIRO.
No mention is made of any of these initiatives when the Report covers Australia’s “Temperate grasslands” in particular the Murray Darling Basin. It states “…These regions are devoid of trees, except for riparian or gallery forests associated with streams and rivers. …and soil Carbon stocks are declining under current management (MacKenzie et al 2017).” Both statements are misleading.
Firstly, while historical land clearing removed trees in some regions of the Basin, much of the land was grassland savanna. Never-the-less the landcare programs launched in 1990 and widely adopted by farmers across the Basin have seen millions of trees and shrubs replanted and the planting is continuing.
Secondly, the reference quoted, MacKenzie et al 2017, for the generalisation that soil carbon stocks are declining misrepresents what the scientists found. This is what the reference said: “… the 60 Priorities for improving soil condition across Australia’s agricultural landscapes results point to a more fundamental set of factors that determine whether soil carbon stocks can be increased:
• Net Primary Productivity needs to be maintained at a high level relative to the potential at the location of interest to ensure that carbon capture by the plant flows through to the soil. In grazing systems this means that overgrazing has to be avoided. In cropping systems, varietal selection, the timing of key operations (e.g. sowing) and application of the right fertiliser at the right time all need to be optimizing to maximize the production of dry matter.
• Water-use has to be efficient and this entails overcoming soil constraints that may be chemical, physical or biological in nature.”
The scientists also noted when discussing increasing soil organic carbon “…that broad patterns of soil carbon variation correlate with first order drivers such as climate and soil type. It also demonstrated that increasing soil stocks in some environments is difficult.”
Given the FAO report is about the state of knowledge of soil biology it is surprising that the authors failed to highlight the pivotal role biota have on the potential for soil to retain soil carbon. The MacKenzie et al 2017 report it uses to make the statement that “…carbon stocks are declining” fails to mention that soil biology itself limits the ability of soils to retain carbon, its why soil carbon stock naturally vary from region to region.
MacKenzie et al 2017, states: “Losses of soil organic carbon due to decomposition are dependent on the activity of the microbial biomass present in the soil. Two key soil properties governing microbial activity are the availability of water and heat. These properties have been mapped and classified into five equal-interval classes for Australia. Class 1 corresponds with environments that have the most rapid decomposition rates and they tend to be hot and dry. Class 5 corresponds with environments that have the lowest decomposition rates (generally the coldest locations although areas that are also very wet may fall into Class 4).”
The figure shows the relationship between soil microbial activity and potential to increase soil carbon across Australia. The approximate area of the Murray Darling Basis in shown and the data demonstrates the variation in soil microbial activity which puts a floor in potential for soils to store carbon.
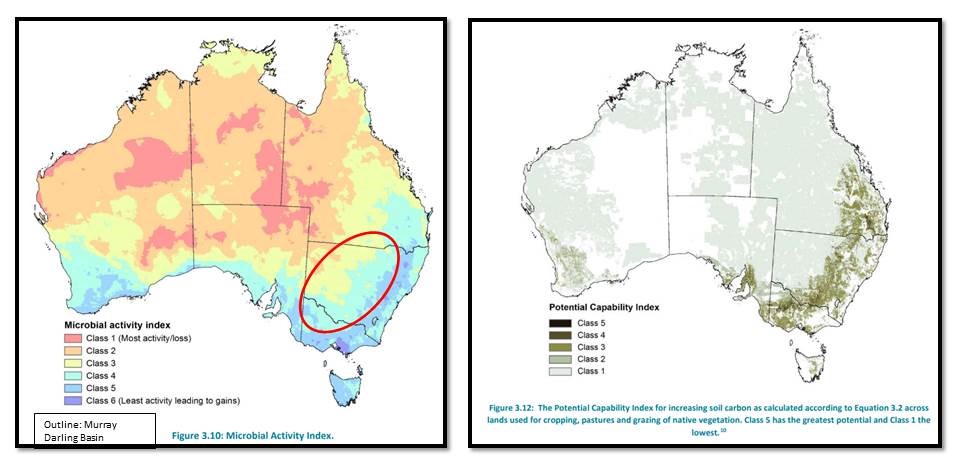
Figure: In contrast to the FAO report which emphasises manipulating soil biota, the MacKenzie et al 2017 review presents opportunities where microbial activity can lead to gains in soil carbon across Australia’s Murray-Darling Basin.
The FAO report’s Chapter five reviews “novel whole-ecosystem approaches for soil restoration and soil management”. It starts by describing the long-held accusations about management of agricultural land that it has reduced soil biodiversity due to tillage, mineral fertilisers and pesticides. But fails to give any recognition in the wide variations in land management around the globe and the wide variation in soils, temperatures and rainfall that are responsible for the microbial activity index as explained in Mackenzie et al 2017. Such lack of context behind accusations blaming agriculture for land degradation is common in FAO reporting.
It seems to misunderstand that soil biomass follows the levels of carbon flows above and in the soil. The more food provided to biota by farming method employed and the environment, the greater the diversity and mass of biota. So fertilisers that promote plant growth (roots and tops) and pesticides which enable more productive species to be planted all increase carbon flows and are recognised in the MacKenzie et al paper.
MacKenzie et al introduces the concept of a “Carbon Gains Index” to describe carbon flows into and out of the soil and so provide food for biota. “The Carbon Gains Index aims to summarize the potential to increase the inputs of organic carbon to the soil. Over the majority of Australia under dryland agriculture, the availability of water places a constraint on productivity which translates into a constraint on organic carbon addition to the soil. The greatest additions of organic carbon in the form of plant residues will occur where transpiration losses of water are maximised and direct evaporation, leaching and runoff losses of water are minimised.
“Potential increases in plant residue inputs under current cropping/pasture/agroforestry systems are possible if one or more of the following conditions exist:
1. Agricultural systems are not using water efficiently and the potential exists to utilise additional water to grow bigger crops (e.g. through choice of cultivars, rotations, nutrient management).
2. Existing constraints to efficient use of plant-available water can be overcome by management (e.g. deep ripping, and mitigating subsoil constraints).
3. Extension of the growing season by inclusion of perennials which will allow carbon to be captured by plants for a greater duration within a year.
4. Crops are grown when the soil contains enough water irrespective of time of year to maximise water use and reduce water lost via evaporation or leaching (e.g. opportunity cropping initiated when the soil profile fills in a manner typical of the northern cereal producing region).”
Instead of explaining these natural factors that influence carbon flow into the soil and hence SOM and SOC for biota diversity and mass, the FAO report concentrates on integrated management of whole soil communities to restore biodiversity and functioning (with the assumption that this is necessary everywhere).
It suggests three novel methods to manipulate and control whole soil biota:
* Whole soil microbiome plus micro-fauna inoculation. Instead of using laboratory bred microbes this process involves spreading donor soil with all its biota diversity onto the recipient “degraded soil”. “However, not all field trials have realized the anticipated results. An important question now is when and where whole-soil biodiversity inocula are effective, and how their consistent effectiveness may be achieved.
* Microbiome engineering. This involves breeding crop varieties in conjunction with known rhizosphere biota. Changes have been achieved with this breeding method and resulted in shifts in flowering time and biomass production in one brassica weed species.
* Ex-situ cultivation of soil microbiomes “Using starter inocula sourced in nature it may be possible to start ex situ soil biodiversity conservation programmes via targeted co-cultivation with the host plant species or indeed whole plant communities.
While the FAO report contends farming with chemicals and artificial fertilisers is not sustainable, and suggests addressing soil organisms and plant microbiomes as the way forward for agricultural sustainability, it concludes biota manipulation is not practical yet.
“Currently there are still rather few products on the market, and generally many products show great effects when tested under highly controlled conditions but fail to show reproducible results under field conditions . The reasons are manifold. On the one hand, applied strains are frequently selected for their plant growth-promoting capacities under laboratory conditions, but do not take into account that any applied strain has to survive in the target environment and establish in a highly competitive (biota) environment. Furthermore, microbial biofertilizers or biopesticides do not behave and perform in the same way as their chemical alternatives; they require suitable formulations and application approaches, which still need to be developed. Last, but not least, understanding the role of soil and plant microbiomes has just started, and novel approaches can be expected to benefit from microbial diversity and activities.”
Take home messages
Despite all the research to date there has been little advance on the two most successful biota plant symbiotic inoculants that have been used by farmers for decades –rhizobia bacteria for plant legumes and mycorrhizae fungi for annual cereals. Science will uncover more roles for particular biota classes but the key message for farmers is managing for increased carbon flows into soil through livestock and crop management and protecting the biota present with ground cover and plant diversity.
,
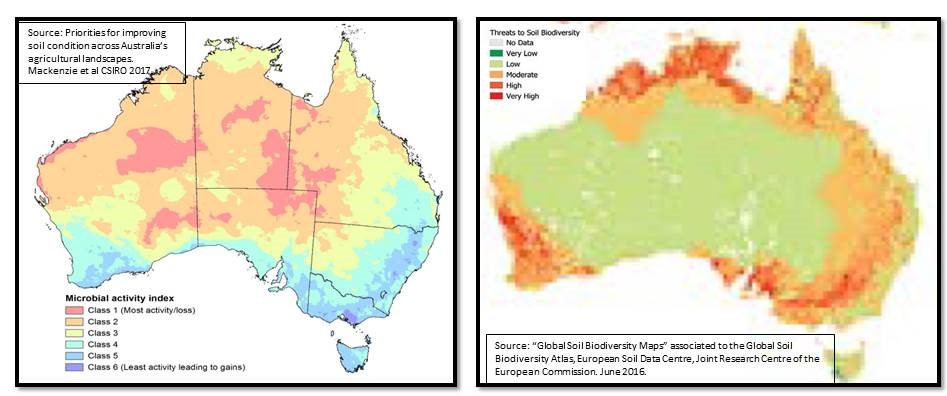
Figure: How two scientific analyses using different parameters for soil biodiversity project different outcomes for soil biology. Left: microbial activity index demonstrates how Australia’s farming regions (blue areas) have high microbial activity which leads to soil carbon gains by increasing carbon flow into the soil through conservation farming methods. Right: Australia’s farming regions are a threat to soil biodiversity and soil carbon. The difference is the MacKenzie et al 2017 scientific papers reflects current and future farming methods while the EU 2016 papers reflects historical impacts of by-gone era farming methods.
Box story
Trophic level definition
A trophic level is the group of organisms within an ecosystem which occupy the same level in a food chain. There are five main trophic levels within a food chain, each of which differs in its nutritional relationship with the primary energy source. The primary energy source in any ecosystem is the Sun (although there are exceptions in deep sea ecosystems).
The solar radiation from the Sun provides the input of energy which is used by primary producers, also known as autotrophs. Primary producers are usually plants and algae, which perform photosynthesis in order to manufacture their own food source. Primary producers make up the first trophic level.
The rest of the trophic levels are made up of consumers, also known as heterotrophs; heterotrophs cannot produce their own food, so must consume other organisms in order to acquire nutrition.
Source: Dictionary of Biology